
Grandes culturas
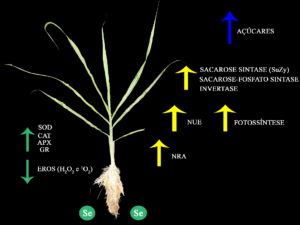
Como aumentar o metabolismo de açúcares e a atividade antioxidante em plantas de cana-de-açúcar?
Figura 1. Ação do selênio (Se) na produção de açúcares a partir do aumento da eficiência do uso no nitrogênio (NUE), taxas fotossintéticas e ativação